





Volume6 No.1
SPring-8 Section A: Scientific Research Report
赤外顕微分光によるアポトーシスを起こした細胞核の解析
Analysis of Apoptotic Nuclei with Infrared Spectra-Microscopy
a量子科学技術研究開発機構, b東北大学, c東京電機大学, d東海大学
aQST, bTohoku University, cTokyo Denki University, dTokai University
- Abstract
-
アポトーシスを誘発したHeLa S3細胞の単離核を赤外顕微分光ビームライン(BL43IR)の試料セルに封入し、赤外顕微分光測定を行い、得られたスペクトルを解析することによりDNAの断片化過程を調べた。アポトーシスを起こしていない正常な細胞核および、アポトーシスの進行を制御した細胞核を用い、リング形状、ネックレス形状、崩壊状態のそれぞれの各ステージにある個々の細胞核の赤外線吸収スペクトルを測定した結果、アポトーシスによる強度の減少が指摘されている 1000 cm-1 付近の PO2- モードのスペクトルにアポトーシスのステージによる特徴的な差異が確認できた。
キーワード: 細胞核、アポトーシス、赤外顕微分光法
背景と研究目的:
アポトーシスは細胞内に生じた異常を取り除く生物の自己防衛機構の一つであり、遺伝子によって支配された能動的な細胞死として注目を集めている。多細胞生物の生体内では、癌化した細胞などの異常細胞はアポトーシスによって除去され、これによりほとんどの腫瘍の成長は未然に防がれる。アポトーシス現象により、細胞は①細胞膜構造変化、②核の凝縮とDNA断片化、③アポトーシス小体の形成の3つの段階の実行過程を経て自死し、染色体異常が起きた細胞ではこの機構により他の正常細胞への悪影響を防ぐ。②の過程がさらに3つの段階に分けられ、ステージ1のDNAを含む染色体繊維が核周辺に集まるリング状構造、染色体繊維が核周辺でビーズ状に変化するステージ2、そしてビーズ状構造が核中心部へ崩落するステージ3と進行することがわかっている[1]。さらに、DNAの電気泳動測定の結果、リング構造からネックレス構造へ形態変化を行う際にDNAの断片化が起こることがわかり、それが最終的な核崩壊を引き起こす原因となると考えられている。このように、これまでの分子生物学的研究により、核の構造変化とDNAの断片化の時間的関係が明らかとなってきたが、そのDNAの断片化が核の構造変化においてどのような位置にあるのかを明らかにするためには、このような状況における構成分子の化学構造に関する情報を取得することが必須と考えられる。核の凝縮過程の構造とDNA断片化との直接の関係を調べるには、電気泳動法では難しく、TUNEL法あるいはコメットアッセイ法が用いられるが、その方法を用いてもDNA断片の大きさ、断端の状態に関する全体的な情報を得ることはできない[2]。我々は、軟X線顕微鏡[3][4]による観察で、核の凝縮過程の詳細な構造変化を観察しているが[5]、この方法でも細胞核の構造変化の詳細はわかるが、その凝縮過程における分子変化の詳細を知ることができない。そこで、分子の振動周波数に起因する赤外線吸収分光スペクトルを測定することによりDNAの断片化の詳細を調べ、アポトーシスの機構の解明に資することを目的とした。
実験:
アポトーシスを誘発した HeLa S3 細胞の単離核を赤外顕微分光ビームライン(BL43IR)の試料セルに封入し、赤外顕微分光測定を行い、得られたスペクトルを解析することによりDNAの断片化過程を調べた。あらかじめ準備したアポトーシス核の溶液(約 10 µL)を赤外顕微分光ビームライン(BL43IR)の液体試料セルに封入し、厚さ 6 μm のスペーサーをはさんだ2枚の BaF2 の窓材の間で保持した。液体試料セルに封入した試料を実体顕微鏡により細胞核の状態と密度を確認後、蛍光顕微鏡により典型的なアポトーシス核を選別し(図1にステージ1の細胞核の場合を例示)、検出器としてMCTを使用した赤外線吸収分光顕微鏡HYPERION2000により計測を行った。細胞核は水に浸した状態で測定する必要があるため、液体セルを使用した。この際、細胞核のサイズとほぼ同じ厚さ 6 μm のスペーサーを使用し、水の吸収が極力小さくなる条件で細胞核のスペクトル測定を行った。顕微鏡の空間分解能は細胞核のサイズにあわせて 5 μm 程度になるようアパーチャーで調整した。測定には細胞膜を破壊し単離した HeLa S3 の細胞核にアポトーシスを誘発した試料を用い、アポトーシスの進行過程の各ステージで止めたものを用意しホルムアルデヒドにより固定した。試料として用いるアポトーシス核は、アポトーシスによる細胞核の構造変化の状態を蛍光顕微鏡により観察できるよう、あらかじめ細胞核内のクロマチンにDAPIを用いて蛍光標識を施した。測定用試料として、ステージ0(正常細胞)、ステージ1(リング形状)、ステージ2(ネックレス形状)、ステージ3(崩壊状態)の4種類を用意した。測定はアポトーシスによる強度の減少が指摘されている[6] PO2- モード(~1235 cm-1、~1085 cm-1)を中心に、600 ~ 4000 cm-1(波数分解能 5 cm-1、積算数 1980 scan)の範囲で行った。アポトーシスを起こしていない正常な細胞核および、アポトーシスの進行を制御した細胞核を用い、リング形状、ネックレス形状、崩壊状態のそれぞれの各ステージにある個々の細胞核の赤外線吸収スペクトルを測定し、各ステージにおけるスペクトルの差異を確認した。
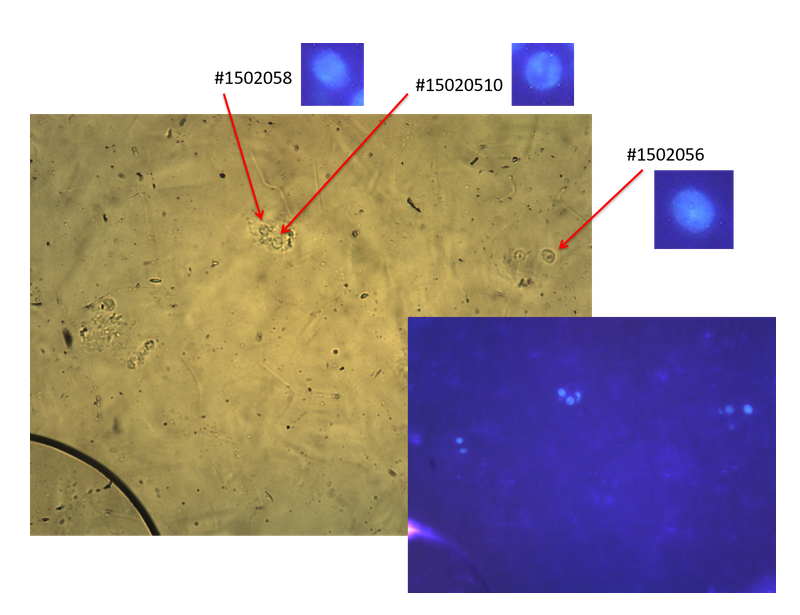
図1. 実体顕微鏡像と蛍光顕微鏡像の比較により典型的なアポトーシスを選別
(ステージ1の細胞核の場合を例示)
結果および考察:
ステージ0(正常細胞)、ステージ1(リング形状)、ステージ2(ネックレス形状)、ステージ3(崩壊状態)における赤外線吸収スペクトルの測定結果を図2に示した。細胞核の個体差による影響を最小化するために各ステージにある複数(3個 ~ 4個)の細胞核の測定結果の平均をとった。あらかじめ実体顕微鏡により典型的な形状の細胞核の選別を行うことにより、スペクトル形状に実験結果の解釈に影響を与えるような大きなばらつきは無かった。アポトーシス過程にある細胞核内で引き起こされているDNAの断片化やタンパク質の構造変化に由来すると考えられる特徴的な 800 cm-1 ~ 1800 cm-1 領域において解析を行った。その結果、1000 cm-1 付近の PO2- に由来する構造の測定と 1600 cm-1 付近のAmideに由来する構造の測定に成功した。特に、1650 cm-1 付近のAmide Iに由来する吸収スペクトルにアポトーシスの各ステージに特徴的な構造が得られた。

図2. 1000 cm-1 付近の PO2- に由来する構造の測定と 1600 cm-1 付近の Amide に由来する構造。特に、1650 cm-1 付近の Amide I に由来する吸収スペクトルにアポトーシスの各ステージに特徴的な構造が得られた。
今後の課題:
図2に示したように、1650 cm-1 付近のAmide Iに由来する吸収スペクトルにアポトーシスの各ステージに特徴的な構造が得られた半面、970 cm-1 付近や 1050 cm-1 付近、850 cm-1 など、ピークの起源を同定出来ていないものも多い。また、ステージ0とステージ1は2回の実験においてよい再現性を見せているが、ステージ2ではピーク a の強度が異なるなど、まだまだ測定の精度や再現性に課題を残している。また、全体的に、バックグラウンドがうまく補正できていない可能性も残している。
参考文献:
[1] S. Tone et al, Exp Cell Res. 313, 3635-3643 (2007).
[2] 竹内隆子 ら、J. Jpn. Soc. Clin. Cytol. 36, 349-357 (1997).
[3] M. Kado et al, Journal of Physics: Conference Series 463, 012056-1-4 (2013).
[4] M. Kado et al, AIP Conference Proceedings 1696, 020019-1-4 (2016).
[5] 加道雅孝 ら、第75回応用物理学会秋季学術講演会、2014年9月17日~20日、北海道大学 札幌キャンパス
[6] B. Mohlenhoff et al, Biophysical Journal 88, 3635-3640 (2005).
ⒸJASRI
(Received: September 4, 2017; Early edition: November 24, 2017; Accepted: December 18, 2017; Published: January 25, 2018)
