





Volume4 No.1
SPring-8 Section B: Industrial Application Report
培養環境におけるコラーゲンゲル構造の品質評価法の開発
Development of the Quality Evaluation Method of Collagen Gel Structure in Culture Environment
日本メナード化粧品(株)総合研究所
Research Laboratories, Nippon Menard Cosmetic Co., Ltd.
- Abstract
-
化粧品の有効性を評価することを目的として、現在様々な試験が行われているが、特に真皮に関しては、コラーゲンゲルを用いた三次元培養系が用いられている。本課題では、より生体に近い培養環境の構築とその評価系の確立につながる情報を得る為に、皮膚組織およびコラーゲンゲルの極小角および小角X線散乱を測定した。その結果、皮膚組織においてコラーゲン線維の径や65 nmの周期に関する構造と思われる回折を捉える事ができた。コラーゲンゲルの回折プロファイルは皮膚組織とは異なるものであったが、対応していると思われる回折がいくつか得られた。また、コラーゲンゲル内のⅢ型コラーゲン比率の変化に伴いゲルの回折が変化することから、コラーゲンの組成によってコラーゲン線維構造が変化していると考えられた。
キーワード: コラーゲンゲル、皮膚、真皮コラーゲン
背景と研究目的:
2009年3月よりEUにおいて化粧品開発に関わる動物実験が禁止されたことを受け、化粧品の有効性を評価するためには、動物に代わり、培養細胞や三次元培養皮膚モデルを用いることが主流になっている。培養細胞は、通常、シャーレ上に平面培養(二次元培養)を行うが、生体環境により近づける為に立体培養(三次元培養)を行うことがある。皮膚においては、コラーゲンゲル(主にⅠ型コラーゲンを用いる)内での真皮線維芽細胞の包埋培養や、さらにその上部に角化細胞を播種・分化させ表皮部位を形成した三次元培養皮膚モデルの構築など、より生体に近い条件においての評価が進められている。しかし、生体反応と異なる結果が得られるケースもあり、より生体に近い評価系が期待されている現状がある。
一方、真皮線維芽細胞は、コラーゲン等の細胞外マトリックスと結合し、情報を認識することで、細胞外環境に依存した変化を示すと考えられている。コラーゲンにはいくつかの種類があり、体の部位によってその構成は異なっている。皮膚の真皮においては、線維成分の90%がコラーゲンで占められており、そのうち約80%がⅠ型コラーゲン、約15%がⅢ型コラーゲンである。これ以外にも、コラーゲン線維間に存在するエラスチン線維や、線維の間隙を満たす基質(プロテオグリカンや糖タンパク質が中心)により、真皮の細胞外環境が形成されている。よって、Ⅰ型のコラーゲンを中心に数種類のコラーゲンやエラスチン、基質などが水を含んだ状態で複雑に組み合わさって存在している生体の真皮中では、これらが形成する微細な構造やその変化が、細胞機能や皮膚性状に影響していると考えられる。しかし、これらの構造や作用メカニズムには、まだまだ不明な点が多い。これまでのコラーゲンゲルを用いた検討において、コラーゲンゲル濃度やコラーゲン組成を変化させると、コラーゲンゲルの弾性やコラーゲンゲルに包埋した線維芽細胞の遺伝子発現が変化することを確認した。これらのことは、コラーゲンゲル構造の変化が物性や細胞機能に作用したと考えている。一方、コラーゲンゲルを走査型電子顕微鏡で観察すると、コラーゲン線維が絡まった構造および線維が束になった構造が観察された(図1)。電子顕微鏡画像から計測したところ、コラーゲンゲル線維の平均直径は49.4 nmであった。電子顕微鏡での観察は、明瞭で分かりやすい画像が得られる利点はあるが、試料を固定・乾燥させる必要があるため、水や基質を含む実際の生体環境とは異なる状態を観察している。
そこで、本課題は、ヒト・ブタ皮膚組織およびコラーゲンゲルの構造を比較するために、固定・乾燥等の前処理を行わず生の状態でX線散乱実験を行うことを目的とした。特に、コラーゲンゲルの線維径を含む微細構造を解析し、さらに組成の異なるコラーゲンゲルの構造変化から、成体に近い培養環境の構築とその評価系について検討した。

図1. コラーゲンゲルの電子顕微鏡画像
実験:
本研究では、市販のコラーゲン溶液(Cellmatrix、新田ゼラチン株式会社製)から作製したコラーゲンゲル、ヒト皮膚組織(TRANSKIN、株式会社ケー・エー・シー製)およびブタ皮膚組織(ユカタンミニブタ皮膚、日本チャールス・リバー株式会社製)を測定用の検体として用いた。組成の異なる3種類のコラーゲンゲルは、市販のⅠ型およびⅢ型のコラーゲン溶液(Cellmatrix type I-AおよびCellmatrix type III)を体積比で10:0、8:2、5:5のそれぞれの比率で混合、pH調整した後、遠心処理によって2倍程度濃縮させたものをゲル試料として用いた。また、ヒトおよびブタの皮膚組織は、真皮部分を2 mm程度の厚さに切り分け、生体とゲルのコラーゲン環境を比較するために使用した。検体は、2枚のカプトン膜に挟んだ後、治具に固定し、試料とした。この2枚のカプトン膜の間隙は2 mmである。なお、各検体は2–3個作製し、データの再現性を確認した。
コラーゲンは、構成単位のトロポコラーゲン(=コラーゲン分子、Ⅰ型コラーゲンの場合、長さ約300 nm、直径約1.5 nm)から始まり、細線維、原線維、線維、線維束という階層構造をとるが、体の部位によってその線維状態が異なる[1]。このように幅広い大きさを持つコラーゲンの構造を捉えるために、極小角X線散乱(USAXS)と小角X線散乱(SAXS)の併用が可能なBL19B2を使用した。測定条件は、X線エネルギーが18 keV、カメラ長がUSAXS 42 m、SAXS 3 m、測定波数(q)レンジはそれぞれUSAXS 0.005–0.2 nm−1、SAXS 0.06–3 nm−1である。カメラ長の較正は、ニワトリの腱由来のコラーゲン線維を用いた。検出器はPILATUS 2Mを使用した。
結果および考察:
ヒト皮膚およびブタ皮膚について、二次元測定データを図2に、これを円環積分し、一次元化したプロファイルおよびそれらの再現性を検討した結果を図3に示す。これら皮膚真皮部位の回折の内、今回は以下の2種類のピークに注目した。
1つ目は、ヒト-ブタ間で共通する位置に存在するピーク群であり、q=0.096 nm−1(面間隔d=65.2 nm)およびq=0.284 nm−1(d=22.1 nm)の位置に見られた(図3左、破線)。トロポコラーゲンが少しずつずれて配向することに起因するコラーゲン細線維の縞模様の周期構造が約67 nmであるので、q=0.096 nm−1(d=65.2 nm)の位置にあるピークはこれを反映している可能性がある。
2つ目は、ヒトとブタで位置が異なるピーク群であり、ヒト皮膚ではq=0.067 nm−1(d=93.2 nm)、q=0.125 nm−1(d=50.1 nm)、q=0.176 nm−1(d=35.7 nm)、q=0.224 nm−1(d=26.9 nm)、ブタ皮膚ではq=0.054 nm−1(d=116.8 nm)、q=0.104 nm−1(d=60.6 nm)、q=0.147 nm−1(d=42.7 nm)、q=0.198 nm−1(d=31.8 nm)のそれぞれ4本が見られた(図3左、実線)。コラーゲン線維は階層的な紐状の構造をしているため、長さと径という2つの構造が考えられる。構成単位であるトロポコラーゲンが長さ約300 nm、直径約1.5 nmとされるので、長さ方向の周期構造である67 nmを除くと、今回観察された300 nm以下の構造は、線維径を反映していると推測された。これらの回折が線維の集合を反映しているとして最密充填を仮定すると、1:1/√3:1/2という比率が当てはまるが、今回観察されたピークの大きさには当てはまらない。したがって、今回観察されたピークは、少なくとも2つ以上の独立した線維径の構造を示していると思われた。ヒト皮膚およびブタ皮膚は、測定する部位により多少のデータのバラツキが見られるが、今回議論しているピークは再現よく観察された(図2右)。
図2. ヒト皮膚およびブタ皮膚の二次元測定データ(SAXS)左:全体図、右:中央拡大図
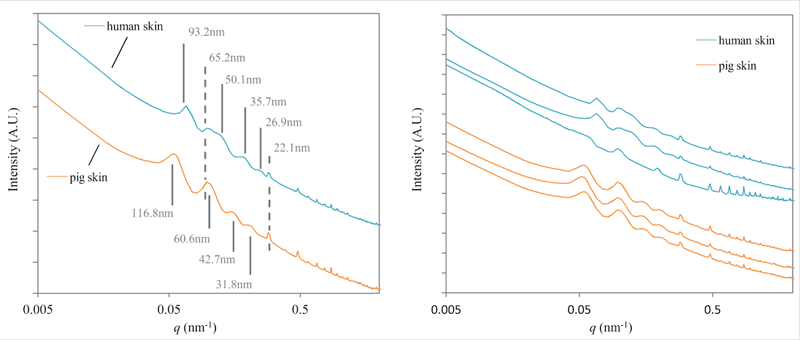
図3. ヒト皮膚およびブタ皮膚の一次元化データおよびそれらの再現性(USAXS + SAXS)
コラーゲンゲルについて、二次元測定データを図4に、これを円環積分し、一次元化したプロファイルおよび再現性を検討した結果を図5に示す。コラーゲンゲルにはブタ由来のコラーゲン溶液を用いているため、今回はブタ皮膚と比較した。コラーゲンゲルのプロファイルは、皮膚組織のプロファイルとは様子が異なっており、q=0.284 nm−1(d=22.1 nm)の小さいピーク(図5左、▲)とq=0.089 nm−1(d=70.5 nm)付近にあるブロードなピーク(図5左、△)が見られた。小さいピーク(▲)はブタの皮膚組織にあるピーク位置と一致する。ブロードなピーク(△)はブタの皮膚組織における4つのピークを含むようなピークと思われる。よって、コラーゲンゲルには皮膚真皮組織と対応すると思われる2種類のピークが観察され、これらは両者に共通する成分であるコラーゲンが作る構造に由来すると考えられた。コラーゲンゲルの電子顕微鏡画像(図1)から算出した平均線維径49.4 nmは、やや小さいがゲルに見られるブロードなピークに含まれることから、観察されたブロードなピークは線維径を反映したものであり、22.1 nmのピークはより小さい単位の線維径であると推測された。加えて、ゲル内のⅢ型コラーゲン比率が増えると、ブロードなピークの中のより大きい成分が増え、ピーク幅がより小角側へ広がることが観察された。
それゆえ、この構造がコラーゲン線維の径を反映していると仮定すると、Ⅲ型コラーゲンを含むことによりコラーゲンゲル線維の平均直径が増加したと考えられた。Ⅰ型コラーゲンから成る線維は分子が密に詰まったものであるのに対し、Ⅲ型コラーゲンから成る線維は分子の詰まり具合が疎であるとされる[2]。これを踏まえると、線維内にⅢ型コラーゲン分子が増えると、分子間に隙間ができ、結果として線維径が増加しているのではないかと考えられた。
図4. ブタ皮膚およびコラーゲンゲルの二次元測定データ(SAXS)左:全体図、右:中央拡大図
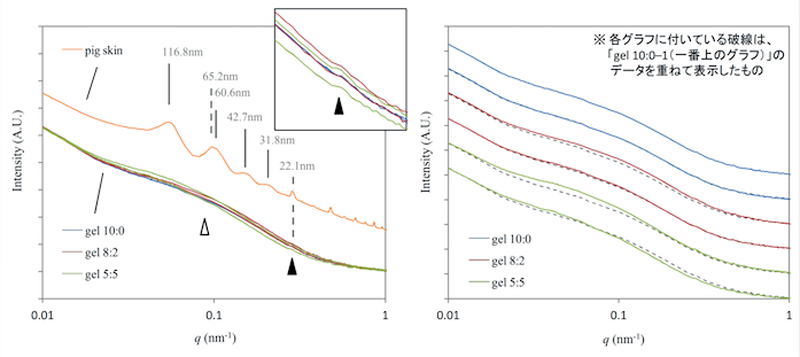
図5. ブタ皮膚およびコラーゲンゲルの一次元化データおよびコラーゲンゲル測定の再現性
(USAXS + SAXS)
今後の課題:
本課題においては、水を抱えた生の状態の試料を用いて、皮膚真皮組織やコラーゲンゲルの回折像を捉える事ができ、両者に対応するような回折も見られた。また、Ⅲ型コラーゲンの比率を変化させることで、その回折にも変化が見られることが確認できた。一方で、組織とゲルの回折像は明らかに異なっていた。その理由として「コラーゲンの結晶化度が低く、ゲルでは明瞭なピークが見えない」ことや「組織のコラーゲンが修飾タンパク質などの作用で一定の大きさの線維に揃っている」ことが考えられたが、詳細については更なる検討を要する。したがって、コラーゲンゲルの評価法として、本手法は培養環境下のコラーゲンゲルと同様に水を抱えた状態のゲルの微細な構造を議論できる点で顕微鏡的手法とは異なる知見を得ることができるものであった。しかし、ゲルを比較・評価していくためには、より明瞭なX線散乱プロファイルが得られるサンプルないし光学系が必要である。
今後は、より顕著な像が見られるようにするための条件検討や今回観察されたピークの帰属を行い、コラーゲンゲルを用いた際に観察できる構造を決定する必要がある。また、Ⅲ型コラーゲンの作用だけでなく、ヒアルロン酸などの基質やエラスチン線維、修飾タンパク質などの影響を確認することで、より生体に近い培養環境が構築でき、細胞外環境の有用な評価系が確立できると考えられる。
参考文献:
[1] Kastelic, J., Galeski, A. and Baer, E., Connective Tissue Research, 6(1), 11-23, (1978).
[2] Junqueira L.C., Montes G.S., Martins J.E., Joazeiro P.P., Histochemistry, 79(3), 397-403, (1983).
ⒸJASRI
(Received: April 9, 2014; Early edition: November 30, 2015; Accepted: December 11, 2015; Published: January 25, 2016)
