





Volume4 No.1
SPring-8 Section B: Industrial Application Report
コラーゲンゲルの品質評価法の開発
Development of the Quality Evaluation Method of the Collagen Gel
日本メナード化粧品(株)総合研究所
Research Laboratories, Nippon Menard Cosmetic Co., Ltd.
- Abstract
-
化粧品の有効性を評価することを目的として利用されるコラーゲンゲルを用いた三次元培養系について、より生体に近い培養環境の構築とその評価系の確立につながる情報を得るために、皮膚組織およびコラーゲンゲルの極小角および小角X線散乱を測定した。その結果、細胞や遠心処理を用いてコラーゲンゲルを濃縮すると散乱強度の増加が認められた。しかし、皮膚組織と対応するような位置にピークが観察されるなどの顕著な波形の変化は見られなかった。また、固定した皮膚組織のうち、メタノール処理した組織では、X線散乱に変化が見られた。すなわち、水を抱えた生の状態の皮膚真皮組織には、メタノール処理により弱まる散乱と強まる散乱があることが分かった。したがって、生の皮膚組織についてX線散乱を測定すると、水環境に依存するような構造が見られると思われた。
キーワード: コラーゲンゲル、皮膚、真皮、線維芽細胞
背景と研究目的:
2009年3月よりEUにおいて化粧品開発に関わる動物実験が禁止されたことを受け、化粧品の有効性を評価するためには、動物に代わり培養細胞を用いることが主流となっている。培養細胞は、通常、シャーレ上に平面培養(二次元培養)を行うが、生体環境により近づけるために立体培養(三次元培養)を行うことがある。特に皮膚分野においては、コラーゲンゲル(主にⅠ型コラーゲンを用いる)内に真皮線維芽細胞を包埋培養した三次元培養系や、さらにその上部に角化細胞を播種・分化させ表皮部位を形成した三次元培養皮膚モデルなど、より生体に近い条件においての評価が進められている。しかし、生体反応と異なる結果が得られるケースもあり、より生体に近い評価系が期待されている現状がある。皮膚の真皮においては、線維成分の90%を占めるコラーゲン線維やコラーゲン線維間に存在するエラスチン線維、線維の間隙を満たす基質(プロテオグリカンや糖タンパク質が中心)により細胞外環境が形成されている。真皮線維芽細胞は、コラーゲンなどの細胞外マトリックスと結合し、情報を認識することで、細胞外環境に依存した変化を示すと考えられている。
よって、コラーゲンやエラスチン、基質などが複雑に組み合わさって存在している生体の真皮中では、これらが形成する微細な構造やその変化が、細胞機能や皮膚性状に影響していると考えられる。しかし、これらの構造や作用メカニズムには、まだまだ不明な点が多い。これまでのコラーゲンゲルを用いた検討において、コラーゲンゲル濃度やコラーゲン組成を変化させると、コラーゲンゲルの弾性やコラーゲンゲルに包埋した線維芽細胞の遺伝子発現が変化することを確認した。また、前回の測定(課題番号2013B1603)においてコラーゲンゲルの線維径に由来すると思われるX線散乱を観察し、コラーゲン組成が線維径を変化させる可能性が示唆された。これらのことは、コラーゲンゲルの構成成分がその構造を変化させ、物性や細胞機能に作用した結果だと考えられる。一方で、実際の皮膚真皮組織とコラーゲンゲルとでは、X線散乱像が明らかに異なっており、それらの構造に明瞭な差があることが確認できた。コラーゲンゲルは、線維芽細胞を包埋培養することにより濃縮される。これは細胞がコラーゲン線維を把握し収縮させることに起因しており、濃縮の程度はゲルに加える細胞数と培養日数に依存する。したがって、コラーゲンゲルのX線散乱が皮膚真皮組織と異なっていたのは、コラーゲンゲルが濃縮されておらず、X線散乱がはっきりと見えなかったためと思われる。
そこで本課題においては、濃縮したコラーゲンゲルに皮膚真皮組織と対応するようなX線散乱が得られるかどうかを検討した。また、水を含んだ生の状態の皮膚真皮組織に特徴的な構造とこれを保持する固定方法を検討するために、組織染色法などで一般的に行われる固定処理により皮膚真皮組織のX線散乱に変化が見られるかを併せて検討した。すなわち、本課題は、濃縮したコラーゲンゲルや固定処理を行った皮膚真皮組織のX線散乱を検討することで、より生体に近い培養環境の構築とその評価系の確立につながる情報を得ることを目的とした。
実験:
本研究では、市販のコラーゲン溶液(Cellmatrix、新田ゼラチン株式会社製)から作製したコラーゲンゲルおよびブタ皮膚組織(ユカタンミニブタ皮膚、日本チャールス・リバー株式会社製)を測定用の検体として用いた。コラーゲンゲルは、線維芽細胞を加えることで収縮させたゲルと遠心処理により濃縮したゲルの2種類をそれぞれ用意した。また、固定処理したブタ皮膚として、メタノールもしくは3%パラホルムアルデヒド/PBSに60分浸したブタ皮膚もそれぞれ用意した。検体は、2枚のカプトン膜に挟んだ後、治具に固定して試料とした。この2枚のカプトン膜の間隙は2 mmである。
コラーゲンは、構成単位のトロポコラーゲン(=コラーゲン分子、Ⅰ型コラーゲンの場合、長さ:約300 nm、直径:約1.5 nm)から始まり、細線維、原線維、線維、線維束という階層構造をとるが、体の部位によってその線維状態が異なる[1]。このように幅広い大きさを持つコラーゲンの構造を捉えるために、極小角X線散乱配置(USAXS配置)と小角X線散乱配置(SAXS配置)の併用が可能なBL19B2を使用した。測定条件は、X線エネルギーが18 keV、カメラ長がUSAXS配置:42 m、SAXS配置:3 m、測定波数(q)レンジがUSAXS配置:0.005−0.2 nm−1、SAXS配置:0.06−3 nm−1、露光時間がUSAXS配置:300秒、SAXS配置:10秒である。カメラ長は、ニワトリ腱由来のコラーゲンの散乱プロファイルを用いて較正した。検出器はPILATUS 2Mを使用した。
結果および考察:
コラーゲンゲルおよびブタの皮膚組織の二次元測定データを図1に、これを円環積分し、一次元化したプロファイルを図2に示す。細胞数や培養日数を変えた細胞包埋コラーゲンゲルのX線散乱を測定したところ、細胞数の増加(1×104 → 5×104 cells/mL、ともに6日培養)や培養日数の増加(3日 → 6日培養、ともに5×104 cells/mL)により、q=0.021−0.536 nm−1のX線散乱強度の増加が認められた。しかし、更なる細胞数の増加(5×104 → 2×105 cells/mL、ともに6日培養)によっては変化がなかったことから、q=0.021−0.536 nm−1のX線散乱強度を増加させるために必要な細胞数には上限値があると考えられた。
一方、遠心処理により物理的に水を吐き出させ濃縮したコラーゲンゲルも、線維芽細胞により濃縮したコラーゲンゲルとほぼ同様のプロファイルを描き、遠心時間の増加(15分 → 30分、ともに15000 rpm)によりq=0.021−0.536 nm−1のX線散乱強度がわずかに増加した。しかし、皮膚組織と対応するような位置にピークが観察されるなどの顕著な波形の変化は、線維芽細胞や遠心処理を用いて濃縮したコラーゲンゲルでは見られなかった。また、線維芽細胞により濃縮したコラーゲンゲルを遠心処理により濃縮したコラーゲンゲルと比較すると、q=0.291−1.346 nm−1にブロードな肩がある点で異なっていた(図2右)。細胞懸濁液(cell suspension)にもq=0.255−1.981 nm−1に肩が見られるため、これは細胞に由来するX線散乱であると考えられた。

図1 コラーゲンゲルおよびブタ皮膚組織の二次元測定データ
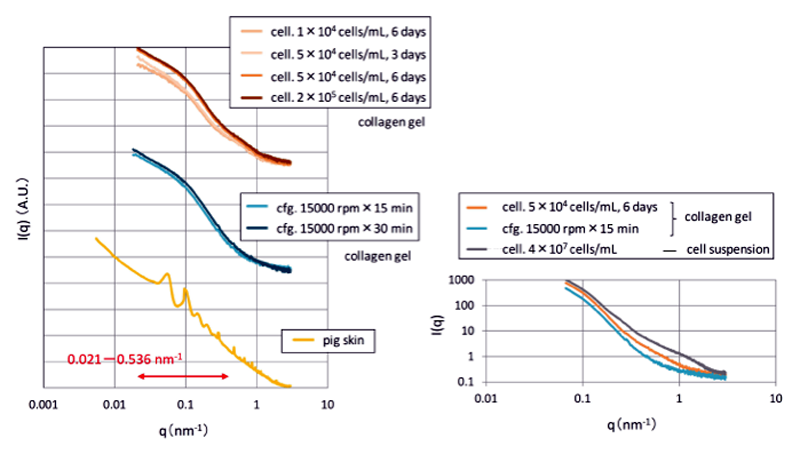
図2 コラーゲンゲルおよびブタ皮膚組織の一次元化データ
メタノール処理およびパラホルムアルデヒド処理は、組織や細胞を固定し、より安定的に組織を保持・観察するために用いられる処理である。今回は、固定処理を行った際にX線散乱で見られる構造が変化しないかを観察するために測定を行った。その結果、パラホルムアルデヒド処理では変化がなかったが、メタノール処理により明らかなX線散乱の変化が見られた。メタノール処理およびパラホルムアルデヒド処理を行ったブタ皮膚組織の二次元測定データを図3に、これを円環積分し、一次元化したプロファイルを図4に示す。
メタノール処理したブタ皮膚では、0.035−0.320 nm−1領域でのピーク群がほとんど見えなくなった一方で、0.385−2.162 nm−1領域でピーク群が顕著になった。よって、水を抱えた生の状態の皮膚真皮組織には、メタノール処理により乱される大きな構造(20−180 nm)と、より結晶化する小さな構造(3−16 nm)があると考えられた。メタノール処理による固定は、脱水作用とタンパク質の凝固作用からなる。それゆえ、メタノール処理により強まる0.385−2.162 nm−1領域のピーク群は、コラーゲン分子の線維径や分子間距離などを含むタンパク質構造に由来すると考えられた。特に、q=0.197 nm−1(2次回折)、q=0.291 nm−1(3次)、q=0.479 nm−1(5次)、q=0.580 nm−1(6次)、q=0.674nm−1(7次)、q=0.768 nm−1(8次)、q=0.869 nm−1(9次)、q=0.963 nm−1(10次)、q=1.057 nm−1(11次)、q=1.158 nm−1(12次)の各ピークは、コラーゲン線維における65 nm周期のブラッグ回折であると考えられた(図4右、▼)。
一方、メタノール処理により弱まる0.035−0.320 nm−1領域のピーク群は、コラーゲン分子が集まってできた構造もしくは他の分子も組み合わさった高次構造に由来すると考えられた。この0.035−0.320 nm−1領域のピーク群は、水を含んだ生の状態の皮膚組織だからこそ観察される真皮の生体構造に由来するのかもしれない。

図3 メタノール処理およびパラホルムアルデヒド処理したブタ皮膚の二次元測定データ
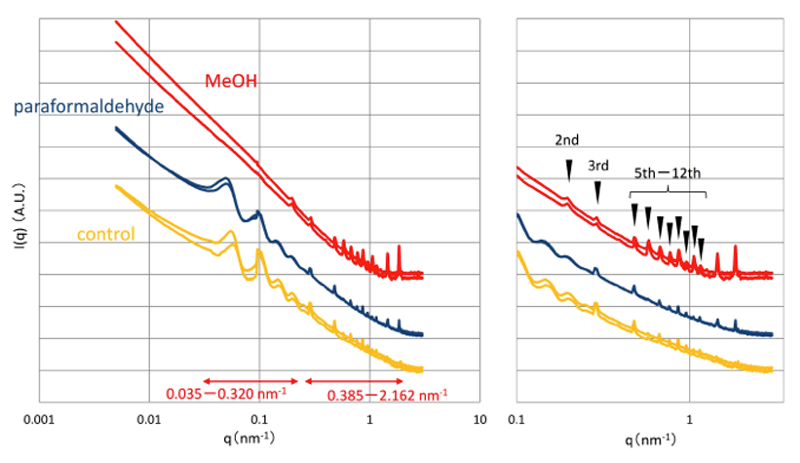
図4 メタノール処理およびパラホルムアルデヒド処理したブタ皮膚の一次元化データ
(左:全体図、右:q > 0.1拡大図)
今後の課題:
本課題において、線維芽細胞や遠心処理を用いたコラーゲンゲルの濃縮によりX線散乱強度の増加が認められた。しかし、この手法でコラーゲンゲルを濃縮しても、これ以上のX線散乱強度の増加は難しいと考えられた。一方、水を含んだ生の状態の皮膚真皮組織から得られるX線散乱が、メタノール処理により変化したことから、生の状態の皮膚を用いると乾燥試料では観察されない水和した構造を見ることができるのかもしれない。
生体内において細胞は、乾燥した細胞外環境ではなく、水和した細胞外環境と相互作用している。それゆえ、より生体に近い培養環境の構築のためには、水和した細胞外環境をいかに再現し、評価できるようにするかが重要である。この水和した細胞外環境は、コラーゲンのみでできているわけではなく、様々な構成成分が関与していると考えられる。例えば、ヒアルロン酸などのムコ多糖は大きな水和構造をとり、プロテオグリカンやコラーゲンと高次構造を作る。今後は、コラーゲンに加え、ヒアルロン酸やプロテオグリカンなどとの相互作用も併せて検討を進め、より生体に近い培養環境の構築および細胞外環境の有用な評価系の確立を目指していく。
参考文献:
[1] Kastelic, J., Galeski, A. and Baer, E., Connective Tissue Research, 6(1), 11−23, (1978).
ⒸJASRI
(Received: August 25, 2014; Early edition: August 25, 2015; Accepted: December 11, 2015; Published: January 25, 2016)
