





Volume8 No.1
SPring-8 Section B: Industrial Application Report
線維芽細胞が産生する真皮線維構造の評価法の開発
Development of an Evaluation Method of Dermal Fiber-like Structures produced by Fibroblasts
日本メナード化粧品(株)総合研究所
Research Laboratories, Nippon Menard Cosmetic Co., Ltd.
- Abstract
-
化粧品の有効性を評価することを目的として、現在様々な試験が行われているが、真皮における「コラーゲンの質」を評価する方法は十分に確立されていない。我々はこれまでに、線維芽細胞の広角・小角X線散乱を測定し、コラーゲンの三重らせん構造、分子間距離、長周期構造に由来する回折について観察してきた。本研究では、線維芽細胞の小角・極小角X線散乱の測定を行い、より大きな構造に由来するX線散乱の観察が可能か検討した。その結果、長周期構造の回折に加え、由来不明の肩ピークが観察された。また、試料の固定、乾燥状態によって、観察されるピークの有無や大小に変化が見られた。
Keywords: コラーゲン、長周期構造、線維芽細胞
背景と研究目的:
コラーゲンは、皮膚や骨、腱などの生体組織の構造を支える主要な線維成分であり、動物の体内に最も多く存在する必要不可欠なタンパク質である。それ故、皮膚や軟骨等を対象とする化粧品や健康食品の開発において、コラーゲンは最も重要な標的物質の一つとされる。また、2009年3月よりEUにおいて化粧品開発に関わる動物実験が禁止されたことを受け、化粧品の有効性評価のためには、動物に代わり培養細胞や三次元培養皮膚モデルを用いた方法が主流となった。このような時代背景の中、培養系を用いたコラーゲンの評価方法をより拡充していくことは、化粧品業界として重要な課題であると考える。
しかし現在のところ、培養系を用いたコラーゲンの評価方法として、「コラーゲンの量」を評価する方法が多用される一方、「コラーゲンの質」を評価する方法は十分に確立されていない。そこで我々は、これまでに本郷らと共に、SPring-8(BL40B2)を利用して、培養細胞が産生する微量なコラーゲンの構造情報をX線散乱により評価するための技術について検討し、三重らせん構造と分子間距離、長周期構造に由来する回折が観察できることを見出した(研究課題 2014B1276、2015A1314、2016A1065、2016B1318)。なお、コラーゲンは階層構造を有し、分子、線維、線維束と様々な大きさの構造を作るが、これまでに培養細胞を用いて観察された構造は分子および線維の構造である(図1)。本研究では、培養細胞として線維芽細胞を用い、本細胞の小角・極小角X線散乱測定を行うことにより、先に記したコラーゲン分子や線維の構造情報に加えて、コラーゲン線維径や線維間距離等のより大きな構造の観察が可能か検討した。
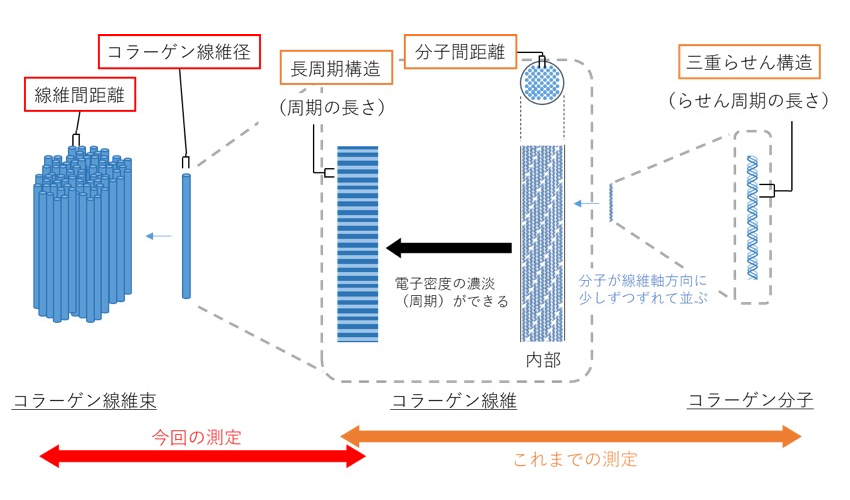
図1. X線散乱測定により観察された培養細胞由来のコラーゲン構造
実験:
線維芽細胞の各試料および比較試料の測定結果について小角・極小角散乱像を円環平均した一次元プロファイルを図2に示した。比較試料であるニワトリ腱由来コラーゲンについては、65 nm の長周期構造に由来する回折が観察された(図2、1–12、数字は回折次数)。一方、線維芽細胞については、明確ではないものの、長周期構造に由来すると思われる回折ピークがどの試料においてもわずかに観察された(図2、矢頭、数字は回折次数)。また、固定による影響について検討するために研究課題 2014A1557 の測定データを引用した。ブタ皮膚試料の小角・極小角散乱像を円環平均した一次元プロファイルを図3に示し、今回のデータを含めて回折について比較した。未処理やパラホルムアルデヒド固定したブタ皮膚とメタノール固定したブタ皮膚とを比較すると、q = 0.55 nm−1 付近を境界にして、長周期構造に由来する回折の見え方が変わった。具体的には、脱水を伴うメタノール固定により、境界より低値の領域にある 65 nm 周期構造由来の1次–5次回折が小さくなる一方、境界より高値の領域にある6次以降の回折が大きく観察された。この特徴は線維芽細胞においても類似したものがあり、脱水されていない未処理およびパラホルムアルデヒド固定の試料においてのみ、3次および5次回折が観察された。複数のピーク位置がほぼ一致しており、メタノール固定による回折の変化の傾向も同じであることから、水和した状態の線維芽細胞の試料において、非常に弱いながらもコラーゲンの長周期構造に由来する回折が観察できると考えられた。また、長周期構造は、メタノール固定に伴う脱水により変化する一方、パラホルムアルデヒド固定では変化しないと考えられた。

図2. 線維芽細胞の小角・極小角散乱像を円環平均した一次元プロファイル
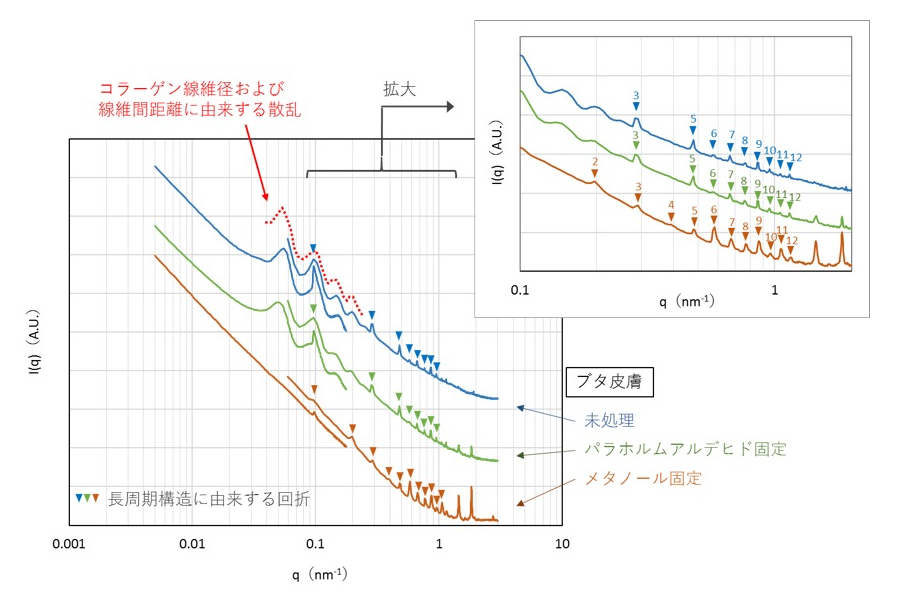
図3. ブタ皮膚の小角・極小角散乱像を円環平均した一次元プロファイル(2014A1557 測定データを引用)
未処理やパラホルムアルデヒド固定を行ったブタ皮膚には、長周期構造に由来する回折に加え、コラーゲン線維径および線維間距離に由来する散乱が q = 0.04–0.25 nm−1 領域に観察された(図3、赤点線)[1]。この散乱は、脱水を伴うメタノール固定によって消失することから、水和した生体構造に由来すると考えられた。一方、線維芽細胞においては q = 0.1 nm−1 付近にブロードな肩ピークが観察され、この散乱は、未処理、パラホルムアルデヒド固定において比較的顕著であった(図2、赤点線)。以前、水和したコラーゲンゲルの測定を行った際にも、線維芽細胞と同じような位置にブロードな肩ピークが観察された[2, 3]。これらは同じ構造に由来する散乱ではないかと考えられた。この肩ピークはメタノール固定および乾燥処理により変化した。具体的には、q = 0.1 nm−1 付近の肩ピークがやや小さくなり、0.3 nm−1 付近にもう一つのブロードな肩ピークが出現した(図2、赤点線)。したがって、この肩ピークは、脱水により変化する構造に由来していると考えられた。本試験では、水和した状態の線維芽細胞の試料において、コラーゲン線維径および線維間距離に由来する散乱は明確に観察できなかった。しかし、似たような位置に水和状態により変化する肩ピークが観察されたことから、これらが同一の構造に由来する散乱である可能性もあると考えられた。
今後の課題:
本研究において、キャピラリに詰めた培養線維芽細胞を用いた場合にでも、長周期構造が観察されることが分かった。また、試料の固定、乾燥状態により、観察されるピークの有無や大小に変化が見られた。特に水和した試料を観察する場合、乾燥した試料やメタノール固定した試料では観察されないコラーゲン線維径や線維間距離といったより大きな構造を観察、評価できるかもしれない。しかし、培養線維芽細胞が産生したコラーゲンを観察する場合、生体組織と比べてコラーゲンが希薄であるためにシグナルが弱く、明確な評価はまだ困難である。この課題を解決するため、今後は、より濃縮した試料形態を検討し、明瞭なピークの観察を目指す。また、本評価法が活用されるためには、構造変化の具体的事例を示すことも必要であると考えられるので、加齢や外部刺激などの皮膚変化に伴うX線散乱像の変化も併せて検証していきたい。
参考文献:
[1] C. Mérigoux et al., ESRF Newsletter 29, 18 (1997).
[2] 足立浩章 他、SPring-8利用研究成果集 4, 93, 2013B1603 (2016).
[3] 足立浩章 他、SPring-8利用研究成果集 4, 111, 2014A1557 (2016).

(Received: October 15, 2018; Early edition: October 30, 2019; Accepted: December 16, 2019; Published: January 22, 2020)
